التهاب نکروتیک روده در جوجههای گوشتی ویژگیهای بیماری و راهکارهای پیشگیری با استفاده از جایگزینهای آلی آنتیبیوتیک

التهاب نکروتیک روده در جوجههای گوشتی: ویژگیهای
بیماری و راهکارهای پیشگیری با استفاده از جایگزینهای آلی آنتیبیوتیک – یک مرور جامع
مقدمه
عفونتهای دستگاه گوارش یکی از نگرانیهای اصلی در صنعت طیور محسوب میشوند و خسارات اقتصادی قابل توجهی را به همراه دارند. انتریت نکروتیک (NE)، چه بهصورت بالینی و چه تحتبالینی، یکی از بیماریهای مهم رودهای در طیور است که تأثیر منفی قابل توجهی بر سودآوری صنعت مرغ گوشتی دارد Bansal et al., 2021;) Salem et al., 2021).
بر اساس یک بررسی انجامشده توسط ون در اسلویس (2000)، تخمین زده میشود که خسارت NE تحتبالینی میتواند تا ۵ سنت به ازای هر پرنده باشد، و شیوع این بیماری ممکن است سالانه حدود ۲ میلیارد دلار برای صنعت جهانی مرغ گوشتی هزینه ایجاد کند. تیمبرمونت و همکاران (2011) نیز خسارت سالانه NE را برای صنعت طیور جهانی تقریباً در همین حدود برآورد کردهاند، که شامل خسارتهای ناشی از کاهش تولید و هزینه های اقدامات کنترلی میشود.
در گذشته، Clostridium perfringensبا نامهای باسیلوس ولچی (Bacillus welchii) و سپس کلستریدیوم ولچی (Clostridium welchii) شناخته میشد، که از ضایعات رودهای پولتهای سیاه اورینگتون در استرالیا جدا شده بود .(Bentz, 1930) بعدها مشاهده شد که این باکتریها مخاط روده پرندگان را آلوده می کنند (Johansson& Sarles, 1928). اصطلاح «انتریت نکروتیک» نخستینبار توسط پریش در سال 1961 در انگلستان مطرح و گزارش شد.
پس از شناسایی بیماری، روشهای مختلفی برای کنترل NE توسعه یافت. در ابتدا، آنتیبیوتیکها برای پیشگیری و درمان NE و همچنین بهعنوان محرک رشد جهت افزایش وزن و بهبود راندمان خوراک مورد استفاده قرار گرفتند، که تصور میشد میزان بروز بیماری را کاهش میدهند (Prescott et al., 1978).
محرکهای رشد آنتیبیوتیکی (AGP) برای دههها در تولید خوراک دام بهطور گسترده مورد استفاده قرار گرفتند (Stokstad & Jukes, 1950). سازمان غذا و داروی ایالات متحده (FDA) استفاده از آنتیبیوتیکها در خوراک دام را در سال 1951 تصویب کرد (Jones & Ricke, 2003). داروهای آنتی بیوتیکی مانند آووپارسین، لینکومایسین، آموکسیسیلین، تایلوزین، ویرجینیامایسین و باسیتراسین معمولاً برای درمان و پیشگیری از NE به کار میرفتند (Krywonos et al., 2001; McDevitt et al., 2006).
با این حال، پس از مشخص شدن نقش AGPها در توسعه و گسترش مقاومت دارویی باکتریها، نگرانیها افزایش یافت و فشار بیشتری برای کاهش استفاده از آنتیبیوتیکها در طیور ایجاد شد. اگرچه تهدید اصلی، افزایش مقاومت آنتیبیوتیکی بود، اما تأثیر حذف داروها نیز قابل چشمپوشی نبود .(Hirshberger et al., 2005)
اتحادیه اروپا در سال 2006
مقررات (EC) شماره 1831/2003 را برای ممنوعیت استفاده از محرکهای رشد آنتی
بیوتیکی در خوراک دام تصویب کرد. سازمان غذا و داروی آمریکا نیز اخیراً اعلام کرده
که قانون «دستورالعمل خوراک دامپزشکی (VFD) را اجرا خواهد کرد (Department
of Health and Human Services, 2015). این قانون تضمین میکند که استفاده
از آنتیبیوتیکهایی که برای سلامت انسان حیاتی هستند، برای اهداف تولیدی ممنوع
بوده و تنها با مجوز دامپزشکی برای درمان بیماریها مجاز است.
در صنعت مرغ گوشتی، حذف AGPها از مزارع تجاری منجر به
افزایش چشمگیر بروز بیماریهای اقتصادی مهمی مانند NE شد (Kysel et al., 2003). برای مثال، پس از حذف آووپارسین در نروژ، موارد NE افزایش یافت. در یک مطالعه، هیچ
کدام از گلههای پرورشیافته در سیستمهای مرسوم با استفاده از آنتی بیوتیک ها مبتلا
به NE نبودند، در حالی که در گلههای
بدون داروی آنتی بیوتیکی، 27 درصد موارد بالینی و 19 درصد موارد تحتبالینی NE
مشاهده شد .(Kacher et al., 2015) این یافتهها
از این دیدگاه حمایت میکنند که حذف افزودنیهای خوراکی میتواند بروز اختلالات
رودهای، بهویژه NE را افزایش دهد .(Sarson et al., 2009)
اگرچه اقدامات سختگیرانه امنیت
زیستی بدون استفاده از AGPها میتواند شیوع بیماری را در برخی مزارع حفظ کند (Engster et al., 2002)، اما اتکا
به این رویکرد برای همه مزارع دشوار است. بنابراین، جایگزینهای AGP برای کنترل NE و سایر بیماریها بسیار مورد
نیاز هستند. در برنامههای بدون دارو، بهجای استفاده از آنتیبیوتیکها و
کوکسیدیواستاتهای خوراکی، از جایگزینهایی مانند روغنهای ضروری استفاده شده است (Krywonos et al., 2004; Zal-Shal et al., 2022).
این مقاله به بررسی عوامل حدت C. perfringens در بیماریزایی NE، واکسنهای موجود، و تأکید بر استفاده از داروهای گیاهی بهعنوان
جایگزین آنتیبیوتیکها برای کنترل و پیشگیری از NE در جوجههای گوشتی میپردازد.
کلستریدیوم
پرفرنژنس و عوامل خطر بیماری
Clostridium perfringens عامل اصلی اختلالات رودهای در حیوانات و انسانها
محسوب میشود Sankar) 1996; Khalifa et al.,
2012b).
این باکتری در محیطهای مختلفی از جمله
فاضلاب، گرد و غبار، هوا و رودههای سالم انسان و حیوان یافت میشود (Khalifa et al., 2012a). در جوجههای گوشتی، بیماری NE معمولاً بین سنین ۲ تا ۶ هفتگی بروز میکند.
این باکتری بیهوازی، گرم مثبت،
اندوسپورساز و غیرمتحرک است و توانایی تشکیل اندوسپورهای مقاوم را دارد که به آن
امکان میدهد در شرایط محیطی سخت مانند مواد آلی در حال فساد و خاک زنده بماند و
پایدار باقی بماند .(Nowak et al., 2003; Khalifa et al., 2015) C. perfringens. قادر به سنتز برخی اسیدهای آمینه ضروری نیست، اما آنزیمهایی
آزاد میکند که بافت میزبان را تخریب کرده و نیازهای تغذیهای خود را تأمین میکند
(Shimizu et al., 2002).
در انسان و حیوانات، توانایی
این باکتری در آزادسازی سموم، رشد سریع در دامنه دمایی گسترده، و تشکیل
اندوسپورها، آن را به یک عامل بیماریزا تبدیل کرده است.
C. perfringens با چسبیدن به پرزهای روده کوچک، تکثیر میشود و سمومی
آزاد میکند که باعث نکروز بافتی شده و در نهایت منجر به بروز NE میگردد
(Shimizu
et al., 2002).
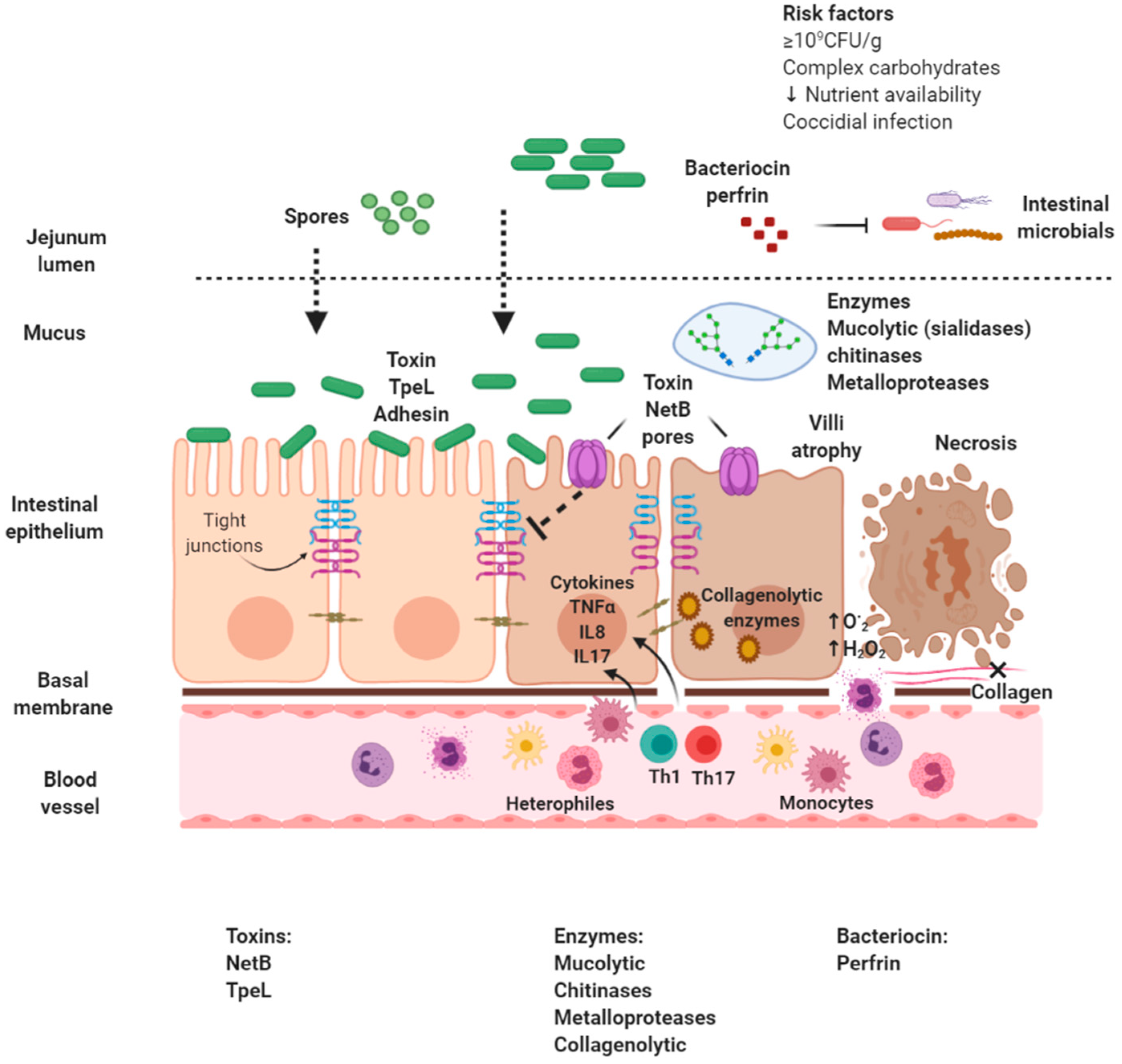
در شکل
یک عوامل مخاطره آمیز یا علل انتریت
نکروتیک با شیوع بالای جهانی بهصورت شماتیک نمایش داده شدهاند.

شکل 1: عوﺍﻣﻞ ﺧﻄﺮ ﯾﺎ ﻋﻠﻞ ﺍﻧﺘﺮﯾﺖ ﻧﮑﺮﻭﺗﯿﮏ ﺑﺎ ﺷﯿﻮﻉ ﺑﺎﻻﯼ
ﻓﻌﻠﯽ ﺩﺭ ﺳﺮﺍﺳﺮ ﺟﻬﺎﻥ
انواع
سموم تولیدشده توسط C. perfringens
سموم تولیدشده توسط Clostridium
perfringens نقش
حیاتی در بروز بیماری انتریت نکروتیک دارند. تاکنون بیش از ۲۰ نوع
مختلف از سموم این باکتری مورد مطالعه قرار گرفتهاند (Lee et
al., 2013). بر اساس وجود ژنهای کدکننده
سموم، C. perfringens
به هفت تیپ (A) تا (G) طبقهبندی میشود. این تیپها
بر اساس نوع سم تولیدی شامل آلفا (α)، بتا (β)، اپسیلون (ε)، یوتا (ι)،
انتروتوکسین (CPE)، بتا۲ (β2) و سمی بنام NetB مشخص میشوند (Sankar, 1996; Opengart, 2020).
هر نوع سم با یک بیماری خاص
انسانی یا حیوانی مرتبط است، که حدت بیماریزایی C.
perfringens به طور مستقیم با تولید این سموم و انتروتوکسینها مرتبط
است (Smedley et al., 2003).
تفاوتهای موجود میان ژنوتیپها
و نوع سموم تولیدی آنها، طیف وسیعی از بیماریهای مرتبط با C. perfringens را در انسان و حیوانات
ایجاد میکند. بهعنوان مثال، تیپ A میتواند باعث قانقاریای گازی در انسان و بیماریهای رودهای در
انسان و حیوانات شود، در حالی که تیپ C قادر است نکروز مخاطی روده کوچک را در حیوانات اهلی و انسانها ایجاد
کند (Petit et al., 1999)
تیپهای A ،C و G بهدلیل ارتباط با بیماریهای طیور، مورد توجه ویژه صنعت مرغداری
قرار دارند .(Opengart,
2020)
در شکل ۲
مقاله، طبقهبندی تیپها و انواع سموم تولیدشده توسط C.
perfringens بهصورت
شماتیک نمایش داده شدهاند.

ﺷﮑﻞ.۲ﻃﺒﻘﻪ ﺑﻨﺪﯼﮐﻠﺴﺘﺮﯾﺪﯾﻮﻡ ﭘﺮﻓﺮﯾﻨﺠﻨﺲﺑﺮ ﺍﺳﺎﺱ ﺗﻮﻟﯿﺪ
ﺍﮔﺰﻭﺗﻮﮐﺴﯿﻦ ﻫﺎﯼ ﺍﺻﻠﯽ
اپیدمیولوژیClostridium perfringens در طیور
با توجه به حضور گسترده این باکتری
در محیط، منبع اصلیC. perfringens محیط
اطراف است Petit et al.,) 1999). انتقال این باکتری معمولاً بهصورت افقی انجام میشود، چرا که
انتقال عمودی بسیار نادر است. بسیار نادر است که مرغها
فاقدC. perfringens در رودههای خود باشند.
تقریباً ۷۵ تا ۹۵ درصد از مرغهای گوشتی این باکتری را
بهعنوان بخشی از فلور طبیعی روده خود دارند .(Svobodova et al., 2007)
آلودگی با C. perfringens میتواند در داخل سالنهای
پرورش مرغ در طول دوره رشد یا از منابع خارجی منشأ بگیرد. مرغها ممکن است از طریق
خوردن بستر آلوده یا نوشیدن آب از منبع آلوده در معرض این پاتوژن قرار گیرند.
حشرات (از جمله سوسکها و مگسها)، کفش کارکنان، خاک اطراف ورودی سالن و آب راکد
خارج از سالن بهعنوان منابع بالقوه آلودگی شناسایی شدهاند.
علاوه بر این، پاتوژنهای مختلف
میتوانند توسط حشرات، پرندگان وحشی و پستانداران که در اطراف سالن مدفوع دفع میکنند،
منتقل شوند. شیوع C. perfringens در محیط ممکن است بسته به شرایط جوجهکشی،
مزرعه، فصل، سن پرندگان و نوع نمونه متفاوت باشد. (Craven
et al., 2001)
علاوه بر این، شدت بروز انتریت
نکروتیک (NE) در
مرغها میتواند بر اساس ترکیب جیره غذایی تغییر کند؛ عواملی همچون وجود گندم و جو
یا پودر ماهی در خوراک، میزان مواد آنتیبیوتیکی و ضدکوکسیدیوزی، و همچنین محتوای
پروتئین حیوانی و کنجاله سویا در این زمینه تأثیرگذار هستند Prescott) و همکاران، 2016). از سوی دیگر، برخی شرایط محیطی نیز میتوانند نقش
مهمی داشته باشند؛ از جمله بستر مرطوب، استفاده از آمونیاک بهعنوان ضدعفونیکننده،
( Hermans , Morgan، 2007)، تراکم بیش از حد طیور و استرس ) Hoerr ، 2010(. رشد بیشازحد کلستریدیوم
پرفرنژنس (C.
perfringens) بهویژه تیپ
A و سموم وابسته به آن در روده باریک،
عامل اصلی این اختلال است که با اسهال ناگهانی و نکروز مخاطی همراه
است (Porter Jr، 1998). بیماری انتریت نکروتیک
(NE) به دو شکل بالینی (کلستریدیوزیس) و تحتبالینی
(کلستریدیوزیس) تقسیمبندی میگردد. جوجههای گوشتی معمولاً حدود ۳ تا ۴ هفتگی پس از خروج از تخم به NE مبتلا میشوند Løvland) ، Kaldhusdal، 2001).
در گلههای گوشتی، نوع بالینی
بیماری میتواند موجب تلفات سریع و ناگهانی شود؛ بهطوریکه در روزهای
پایانی دوره پرورش، تلفات روزانه به حدود ۱ درصد در روز و بهصورت چند روز متوالی میرسد (Kaldhusdal و Løvland، 2000). علائم
بالینی کلستریدیوزیس شامل کمآبی بدن، افسردگی، اسهال کفآلود به رنگ نارنجی
(که در برخی موارد با خون آمیخته است) و پرهای ژولیده میباشد. در
کالبدگشایی، بخشهای وسیعی از روده دچار نکروز شده و توسط یک غشای کاذب
زرد ـ قهوهای پوشیده میشود که حاوی سلولهای نکروتیک، کلنیهای باکتریایی و
قطعات بافتی است (Lee و همکاران، 2011؛ Timbermont و همکاران، 2011).
اشکال تحتبالینی بیماری تشخیص
دشوارتری دارند و با وجود عدم تلفات آشکار، اثرات اقتصادی بیشتری نسبت به
نوع بالینی ایجاد میکنند ( Stutzو Lawton، 1984 .(نشانههای
کلستریدیوزیس در این حالت بسیار محدود و سختقابلتشخیصاند و ممکن است شامل کاهش
جذب و هضم مواد مغذی، افت ضریب تبدیل خوراک (FCR) و کاهش افزایش وزن زنده (BWG) باشند (Stutz و Lawton، 1984(.
علاوه بر این، وجود ضایعات
نکروتیک خفیف تا متوسط در روده باریک از جمله شاخصهای ابتلای تحتبالینی به
انتریت نکروتیک (NE) محسوب میشود. تشخیص قطعی بیماری از طریق آنالیز
باکتریشناسی و ژنوتایپینگ جدایهها امکانپذیر است (Kaldhusdal و Hofshagen، 1992). همچنین، NE تحتبالینی میتواند با تخریب کیفیت بستر
همراه باشد که این امر خطر ابتلا به درماتیت کف پا و سوختگی مفصل خرگوشی (hock burn) را
افزایش میدهد؛ دو عارضهای که از مشکلات مهم رفاه حیوان در صنعت طیور به شمار میروند (
Allain
و همکاران، 2009).
در بیماری کلستریدیوزیس،
باکتریها میتوانند از طریق مجرای صفراوی به کبد منتقل شده و در آنجا
کلونیزه شوند و منجر به بروز کلانژیوهپاتیت و آسیت (تجمع مایع در
حفره صفاقی) گردند (Kaldhusdal و همکاران، 2001). پرندگانی
که در کبد آنها ضایعات مرتبط با C. perfringens مشاهده
میشود، در کشتارگاه ممکن است ضبط و حذف شوند (Løvland و Kaldhusdal، 1999).
از نظر بافتشناسی، در پرندگان
مبتلا به NE تحتبالینی (در شرایط آزمایشی)، لایه لامینا پروپریا
روده دچار هایپرمی میشود، در حالی که اپیتلیوم نسبتاً طبیعی باقی میماند.
در این شرایط، لنفوسیتها، گرانولوسیتها، سلولهای پلاسما، ماکروفاژها و
تعدادی ائوزینوفیل به لامینا پروپریا نفوذ میکنند. در محل تماس، انتروسیتها
و لامینا پروپریا دچار ادم میشوند. پرزها (ویلی) کوتاه شده و کریپتها
متسع میگردند. نکروز سلولهای اپیتلیال با متراکم شدن کروماتین، کاریورکسیس (به معنی تجزیه و خرد
شدن هسته سلول است.) و کاریولیزیس (به معنی حل شدن و از بین رفتن هسته
است) مشخص میشود (Engberg و همکاران، 2002).
از نظر اقتصادی، خسارت های
بیماری عمدتاً ناشی از افزایش مصرف خوراک به دلیل کاهش کارایی تغذیهای، حذف
پرندگانی که در کشتارگاه ضبط میشوند و همچنین درمانهای مورد نیاز برای
بازگرداندن سلامت گله است. با وجود اینکه پاتوژنز بیماری بهطور کامل شناخته
نشده، شواهد نشان میدهند که آنتیژنها و سموم C. Perfringens به تنهایی قادر به ایجاد بیماری نیستند، مگر در حضور عوامل
مستعدکننده (Cooper و Songer، 2009).
عوامل مستعدکننده و پاتوژنز NE در شکل ۳ نشان داده شدهاند.

شکل ۳. پاتوژنز انتریت نکروتیک
عوامل
مستعدکننده (Predisposing factors)
مراحل
پاتوژنز (شمارههای روی شکل)
1)
ایجاد محیط مناسب برای تکثیر C. perfringens مثل افزایش تولید موکوس
(mucogenesis).
2)
تخریب سلولهای اپیتلیال روده توسط عوامل مستعدکننده (مثل کوکسیدیوز
یا مایکوتوکسینها).
3)
نشت پروتئینهای پلاسما به درون لومن روده.
4)
چسبیدن C. perfringens به اپیتلیوم روده.
5)
ترشح پپتیدهای میکروبی مثل باکتریوسینها و ایجاد دیسباکتریوزیس.
6)
ترشح اگزوتوکسینها مثل آلفا، بتا و NetB
toxin.
7)
نفوذ باکتری از طریق جریان خون باب کبدی (portal
circulation) به کبد و ایجاد کلانژیوهپاتیت
و آسیت.
عفونت
کوکسیدیایی
کوکسیدیا یک انگل تکیاختهای
است که درون روده میزبان تکثیر یافته و موجب بروز بیماریهای گوارشی میشود.
تاکنون هفت گونه از جنس Eimeria شامل E. acervulina، E. maxima، E. mitis، E. praecox، E. necatrix، E. tenella و E. brunetti بهعنوان عوامل بیماریزا در پرندگان شناسایی شدهاند (Williams، 2005). از
میان این گونهها، E. acervulina و E. maxima بهعنوان مهمترین عوامل مستعدکننده ابتلا به انتریت نکروتیک (NE) در مرغها شناخته میشوند
(Al-Sheikhly و Al-Saieg، 1980).
اگرچه کوکسیدیوز و نكروز
انتروتوكسیك (NE) با هم ارتباط دارند و علائم مشابهی نشان میدهند، معمولاً کوکسیدیوز
قبل از شروع یا در طول فاز NE بروز میکند. علاوه بر این، مطالعات نشان دادهاند که استفاده از
واکسن ایمریا (Eimeria) میتواند
شیوع NE را افزایش دهد (Pedersen و همکاران، ۲۰۰۸).
مکانیزم دقیقی که نشان دهد
کوکسیدیا چگونه NE را ایجاد میکند، مشخص نیست و علل احتمالی مختلفی مطرح شدهاند. آسیب
وارد شده به لومن روده، معمولاً در زمان تکثیر کوکسیدیا، منجر به خونریزی میشود؛
در نتیجه پروتئینهای پلاسما که نشت کردهاند، منبعی از بستر رشد برای C. perfringens فراهم میآورند. علاوه بر
این، کوکسیدیوز باعث تحریک تولید مخاط شده و محیط مناسبی برای رشد C. perfringens ایجاد میکند Collier) و همکاران، ۲۰۰۸؛ Kaldhusdal و همکاران، ۲۰۲۱). دادههای توالییابی ژنومی نشان میدهند که C. perfringens قادر
به تولید آنزیمهای مورد نیاز برای سنتز اسیدهای آمینه ضروری نیست (Shimizu و
همکاران، ۲۰۰۲).
Baba و همکاران (۱۹۹۲) نشان دادند که احتمال ماندگاری C.
perfringens در مخاط سکوم جوجههایی که به Eimeria آلوده نشدهاند،
کمتر از جوجههای آلوده است. همزمانی عفونت با E. maxima معمولاً با بروز NE در جوجههای گوشتی همراه است و به همین دلیل در مدلهای
آزمایشگاهی NE به همراه C. perfringens برای ایجاد ضایعات NE به کار میرود، چرا که موجب تغییر ترکیب میکروبیوتای
روده میشود (Wu و همکاران، ۲۰۱۴) E. maxima یک انگل پروتوزوآیی داخلسلولی است که کوکسیدیوز
ایجاد میکند؛ این انگل طی مراحل داخلسلولی چرخه زندگی خود، باعث ایجاد ضایعات
رودهای و تخریب اپیتلیوم روده میگردد. در نتیجه، پروتئینهای پلاسما به درون
لومن نشت کرده و بهعنوان منبع تغذیه برای C. perfringens عمل میکنند.
مشخص شده است که در حیوانات
آلوده به E. maxima، سطح پپتید ضد میکروبی شماره ۲ با
منشأ کبدی کاهش مییابد. این موضوع میتواند شرایط را برای تکثیر C. perfringens ـ عامل ایجادکننده NE ـ در رودهای که قبلاً دچار آسیب شده است، فراهم کند. در یک مدل آزمایشی NE با همزمانی آلودگی به Eimeria و C. perfringens، مشاهده شد
که سایتوکاین سرکوبگر ایمنی، اینترلوکین-۱۰ (IL-10)، بهطور معنیداری افزایش مییابد؛ در حالی که در آلودگی منفرد با
C. perfringens
تغییری مشاهده نشد. این یافته نشان میدهد که حضور انگل میتواند
فرصت بیشتری برای تکثیرC. perfringens فراهم سازد
(Park و همکاران، ۲۰۰۸). سایر
میانجیهای ایمنی همچون IFN-α، IFN-γ و IL-1β در مدلهای آلودگی همزمان، در مقایسه با آلودگی منفرد C. perfringens، طی روزهای
پس از عفونت کاهش بیان داشتند؛ در حالی که IL-8 همزمان با IL-10 افزایش بیان نشان داد
(Parvizi و همکاران، ۲۰۱۰). همچنین در جوجههای گوشتی آلوده
به Eimeria
با دوز ۱۰ برابر واکسن
(Paracox) و C. perfringens که بهطور آزاد تغذیه شدند، شدت
ضایعات رودهای بالاتر از پرندگانی بود که بر اساس پروتکل عفونت، بهمدت ۱۲ ساعت در شب محدودیت غذایی داشتند
(Tsiouris و همکاران، ۲۰۱۴).
عوامل تغذیهای
در طیور، ترکیب جیره غذایی نقش
مهمی در بروز پاتوژنز NE دارد. جیرههایی که حاوی مقادیر بالای مواد غیرقابلهضم و پلیساکاریدهای
غیرنشاستهای (NSP) هستند ـ مانند گندم، جو، چاودار و یولاف ـ بهعنوان عوامل مستعدکننده
برای بروز NE در پرندگان شناخته شدهاند
(Kaldhusdal و همکاران، ۱۹۹۹). جوجههای گوشتی که با جیره
مبتنی بر گندم یا جیره گندمی همراه با کربوهیدراتهای پیچیده و فیبر اضافی تغذیه
شدند، ضایعات رودهای بیشتری نسبت به پرندگانی که جیره بر پایه ذرت دریافت کردند،
نشان دادند (Annett و همکاران، ۲۰۰۲). این موضوع احتمالاً به وجود آرابینوزیلانها و β-گلوکانها مرتبط است؛ ترکیباتی
که دستگاه گوارش طیور قادر به هضم کامل آنها نیست، اما میتوانند بهعنوان بستر
رشد برای میکروبیوتا و همچنین باکتریهای بیماریزا مانند C. perfringens مورد استفاده قرار گیرند
( Annett و همکاران، ۲۰۰۲).
گنجاندن مقادیر بالای غلات در
جیره، که اغلب سرشار از پلیساکاریدهای غیرنشاستهای محلول در آب (NSP) هستند، یکی
از عوامل مستعدکننده برای بروز NE محسوب میشود. افزایش غلظت کربوکسیمتیلسلولز، بهعنوان یک NSP، باعث کاهش
سرعت عبور مواد غذایی از دستگاه گوارش و در نتیجه افزایش ویسکوزیته روده میگردد؛
شرایطی که محیطی بسیار مناسب برای عفونت کلستریدیایی فراهم میآوردTimbermont) و همکاران، ۲۰۱۱). همچنین حضور پروتئینهای هضمنشده در بخش انتهایی روده
جوجههای گوشتی با بروز NE مرتبط دانسته شده است. میزان پروتئین موجود در جیره و ماهیت آن نیز
در ارتباط با شیوع NE گزارش شده است Fernando) و همکاران، ۲۰۱۱).
Fernando و همکاران (۲۰۱۱) نشان دادند که پرندگانی که با جیره حاوی پروتئین سیبزمینی (که
قابلیت هضم پایینی دارد) تغذیه شدند، تیتر بالاتری از آنتیبادی آلفا-توکسین تولید
کرده و افزایش معنیداری در ضایعات کبدی و بروز نکروز رودهای نسبت به پرندگانی که
با پروتئین سویا تغذیه شدند، نشان دادند. همچنین مصرف پروتئینهای با منشأ حیوانی
بهعنوان عامل مستعدکننده NE مطرح شده است. شمار C. perfringens در ایلئوم و سکوم جوجههای گوشتی تغذیهشده با
جیرههای حاوی پودر ماهی یا جیرههای بر پایه پودر گوشت/استخوان بالاتر بود و با
افزایش سطح پروتئین خام جیره نیز افزایش یافت
(Williams، ۲۰۰۵). پودر ماهی منبع غنی از روی،
گلیسین و متیونین است (Dahiya و همکاران، ۲۰۰۶).
حضور پروتئینهای حیوانی میتواند
سبب تحریک سنگدان و افزایش pH آن شود که این شرایط، محیط مطلوبی برای رشد C.
perfringens فراهم میآورد. همچنین چربیهای حیوانی مانند
پیه یا دنبه در مقایسه با روغنهای گیاهی، شمار C.
perfringens در ایلئوم را بیشتر افزایش میدهند (Knarreborg و همکاران، ۲۰۰۲). کیفیت پروتئین موجود در این نوع
جیرهها بالاست، اما همواره بهطور کامل قابل هضم نیست و بخشی از آن به بخشهای
انتهایی دستگاه گوارش عبور میکند. منابع مختلف پروتئینی دارای غلظتهای متفاوتی
از اسیدهای آمینه هستند و برخی از این اسیدهای آمینه میتوانند رشد C. perfringens را تسریع کرده و در نتیجه
در بروز NE نقش داشته باشند. افزایش سطح گلیسین در جیره جوجههای گوشتی باعث
افزایش شمار C. perfringens در
ایلئوم و سکوم میشود. همچنین تغذیه با پروتئین منبع ماهی میتواند پویایی
میکروبیوتای روده جوجهها را تغییر داده و شرایط مناسبی برای کلونیزاسیون C. perfringens در روده فراهم سازد (Wu و
همکاران، ۲۰۱۴)
عوامل
ضدتغذیهای مانند مهارکنندههای پروتئاز،
لکتینها و تاننها میتوانند حساسیت طیور به NE را افزایش دهند. آنتی تریپسین که بهطور معمول در کنجاله
سویا وجود دارند، با باقیگذاشتن پروتئینهای هضمنشده در بخش انتهایی روده، محیطی
ایدهآل برای تکثیر باکتریها فراهم میکنند
(Clarke و
Wiseman، ۲۰۰۷). علاوه بر ماهیت شیمیایی جیره،
حضور میکروارگانیسمهای رودهای میتواند به ماهیت فیزیکی خوراک نیز وابسته باشد (Smith و Macfarlane، ۱۹۹۸). استفاده از آسیاب غلتکی برای خردکردن گندم نشان داده است
که مرگومیر ناشی از NE را در مقایسه با آسیاب چکشی کاهش میدهد. همچنین جیرههای آردی درشتدانه میتوانند ترشح اسیدکلریدریک را تحریک کرده و زمان
ماندگاری خوراک در سنگدان و پیشمعده را افزایش دهند Engberg)
و همکاران، ۲۰۰۴).
علاوه بر این، شکل فیزیکی جیره
ارائهشده به جوجههای گوشتی نیز در رشد پاتوژنز NE نقش دارد. Engberg و همکاران (۲۰۰۲) گزارش کردند که شمار C. perfringens در جوجههای تغذیهشده با
جیره پلت کمتر از جوجههایی بود که جیره مش دریافت کردند، هرچند در این مطالعه
ضایعات رودهای ارزیابی نشد. همچنین در جوجههای گوشتی آلوده به C. perfringens، محدودیت
تغذیهای موجب کاهش معنیدار pH روده و شمار C. perfringens در سکوم (تغییر اکوسیستم رودهای) در مقایسه با گروه
تغذیه آزاد (ad libitum) شد. مکانیسمهای کاهش شدت ضایعات در پرندگان با تغذیه محدود بهطور
کامل مشخص نبود، اما این فرضیه مطرح شد که افزایش سطوح گلوکوکورتیکوئید ناشی از
محدودیت غذایی میتواند منجر به کاهش پروستاگلاندینها و در نتیجه سرکوب التهاب
گردد (Tsiouris و
همکاران، ۲۰۱۴). در یک گزارش اخیر مشخص شد که افزایش وزن بدن (BW) و افزایش
وزن روزانه (BWG) در جوجههای گوشتی میتواند بهعنوان یک عامل مستعدکننده در بروز NE مطرح باشد. از آنجا که نتایج
این مطالعه ارتباطی بین BWG و ایجاد ضایعات NE نشان داد، پژوهشگران بررسی کردند که آیا کاهش BWG (در
مراحل خاصی از چرخه رشد) میتواند بهعنوان بخشی از یک راهکار پیشگیرانه در برابر NE مورد استفاده قرار گیرد (Dierick و
همکاران، ۲۰۱۹).
استرس و سرکوب ایمنی
استرس یکی از عوامل مهم
مستعدکننده ابتلا به انتریت نکروتیک (NE) به شمار میآید. دلیل این موضوع میتواند تغییرات
ایجادشده در دستگاه گوارش باشد (McDevitt و همکاران، 2006). عواملی
مانند حضور آمونیاک در محیط، استرسهای فیزیولوژیک، تراکم بالای پرنده در سالن،
استرس گرمایی و مسمومیت ناشی از مایکوتوکسینها نشان دادهاند که میتوانند سیستم
ایمنی را تضعیف کرده و زمینه را برای ابتلا به NE فراهم کنند (Tsiouris و
همکاران، 2015). همچنین مشخص شده است که استرسهای محیطی مثل استرس سرمایی هم میتوانند
با تضعیف ایمنی باعث بروز
NE شوند. به همین شکل، Burkholder و همکاران (2008) گزارش
کردهاند که استرس گرمایی با تغییر در ساختار روده و برهم زدن تعادل میکروبی آن،
بهعنوان یک عامل مستعدکننده قبل از بروز عفونت عمل میکند (Lee و همکاران، 2011).
یک بررسی در مزارع گوشتی انگلستان
نشان داد که مزارعی با بستر خیس، یا آنهایی که از آمونیاک بهعنوان ماده ضدعفونیکننده
استفاده میکردند، شیوع بالاتری از انتریت نکروتیک (NE) داشتند (Hermans و Morgan، 2007). همچنین تراکم بالای پرنده (ازدحام) بهعنوان یک عامل مستعدکننده مطرح
شده که با کاهش رفاه طیور و آسیب به سلامت روده، شرایط را برای بروز NE فراهم میکند (Tsiouris و همکاران، 2015). علاوه
بر این، بیماریهای سرکوبکننده ایمنی مانند بیماری بورس عفونی، ویروس کمخونی
عفونی جوجهها (CIAV) و بیماری مارک، در تشدید روند ابتلا به NE ناشی از کلستریدیوم پرفرنژنس مؤثر دانسته شدهاند (Lee و
همکاران، 2011).
ویروس بیماری مارک با تخریب
سلولهای B و دگرگونی سلولهای T
باعث بروز سرکوب ایمنی و ایجاد لنفومها میشود (Müller و
همکاران، 2003). ویروس بیماری بورس عفونی نیز سلولهای لنفاوی موجود در بورس فابریسیوس
را هدف قرار داده و در نتیجه، باعث کاهش جمعیت سلولهای لنفاوی و تضعیف سیستم
ایمنی در پرندگان میگردد و به این ترتیب، خطر ابتلا به سایر عفونتها افزایش مییابد (Hailemariam و
همکاران، 2008). کمخونی، آپلازی مغز استخوان (آپلازی مغز استخوان به حالتی گفته میشود
که در آن فعالیت طبیعی مغز استخوان برای تولید سلولهای خونی (شامل گلبولهای
قرمز، گلبولهای سفید و پلاکتها) کاهش یافته یا کاملاً متوقف میشود)، آتروفی
تیموس و سرکوب ایمنی از ویژگیهای بارز ویروس کمخونی عفونی جوجهها (CIAV) بوده و این
عوامل در افزایش بروز
NE نقش دارند (Flores-Diaz و
همکاران، 2005).
پاسخ
ایمنی جوجههای گوشتی در برابر انتریت نکروتیک (NE)
تا مدتها تصور میشد که
استفاده از آنتیبیوتیکهای خوراکی و داروهای ضدکوکسیدیوز مانع از تبدیل NE به یک مشکل جدی در صنعت طیور شده است؛ به همین دلیل پاسخ ایمنی جوجهها
در برابر کلستریدیوم پرفرنژنس چندان مورد مطالعه و شناسایی دقیق قرار
نگرفته بود (شکل 4). مشابه پستانداران، جوجهها نیز دارای دو بخش اصلی سیستم
ایمنی، یعنی ایمنی ذاتی و اکتسابی هستند؛ با این حال، تفاوتهایی بین سیستم ایمنی
پرندگان و پستانداران وجود دارد (Salem وAttia ، 2021).
بهطور کلی، سیستم ایمنی
پرندگان به نظر میرسد گیرندهها و مولکولهای مؤثر کمتری نسبت به پستانداران
داشته باشد. ایمنی ذاتی نخستین خط دفاعی در برابر پاتوژنها را تشکیل میدهد و
شامل گیرندههای شناسایی آنتیژن، سلولهای فاگوسیتی و مولکولهای ترشحی سدّی است (Müller و
همکاران، 2003). در مقابل، پاسخ ایمنی اکتسابی
اختصاصیِ پاتوژن بوده و توسط سلولهای T و B میانجیگری میشود.

شکل 4. پاسخ ایمنی جوجهها به عفونت ناشی از Clostridium perfringens
در ارتباط با NE، بیشترین اهمیت به سیستم ایمنی موجود در روده معطوف است. دستگاه
گوارش جوجه در چند روز پایانی قبل از خروج از تخم و بلافاصله پس از هچ بهسرعت رشد
کرده و برای هضم و جذب مواد مغذی تکامل مییابد (Hailemariam و
همکاران، 2008).
پس از هچ شدن، دستگاه گوارش
جوجهها بهسرعت توسط باکتریها کلونیزه میشود و باید میان دو وظیفه اصلی خود
یعنی هضم مواد غذایی و محافظت میزبان در برابر پاتوژنها تعادل برقرار کند. نخستین
لایههای محافظتی شامل موانع فیزیکی هستند که از اتصال پاتوژنها به سلولهای اپیتلیال
جلوگیری میکنند و همچنین موانع شیمیایی که میتوانند غشای سلولهای میکروبی را
تخریب نمایند
(Tsiouris و همکاران، 2015).
کلونیزاسیون میکروبی دستگاه
گوارش در پرندگان برای تکامل سیستم ایمنی ضروری است. پرندگانی که در شرایط عاری از
میکروب نگهداری میشوند، در مقایسه با پرندگان پرورشیافته در شرایط معمولی،
فولیکولهای لنفاوی ضعیفتری در ناحیه لوزه سکومی دارند و در سن ۴ هفتگی هیچ سلول مثبت برای IgG یا IgA در آنها شناسایی نمیشود.
همچنین در پرندگان عاری از میکروب، شمار سلولهای
(CD3+) T در ناحیه پرزهای سکومی کمتر از پرندگان معمولی گزارش شده
است Honjo) و همکاران، 1993).
بیان ژن CD3 که نشانگر حضور سلولهای T است،
در روزهای ابتدایی زندگی در سطح پایین قرار دارد اما از روز چهارم به بعد بهطور
قابل توجهی افزایش مییابد که بیانگر رشد جمعیت سلولهای T در روده طی گذر زمان است (Bar-Shira و همکاران، 2003). سیستم
ایمنی روده باید توانایی تمایز میان باکتریهای همزیست و باکتریهای بیماریزا را
بهدست آورد تا پاسخهای مؤثر تنها علیه ارگانیسمهایی فعال شود که قابلیت تهاجم و
تخریب بافتهای میزبان را دارند (Bar-Shira و همکاران، 2003).
پاسخهای
ذاتی (Innate Responses)
مولکولهای
سدّی (Barrier
Molecules)
یکی از نخستین خطوط دفاعی در
روده جوجهها، لایه مخاطی (mucin
barrier) است
و این مولکولهای موکوس احتمالاً از اولین مولکولهایی هستند که با perfringens Clostridium
مواجه
میشوند. در ژنوم جوجهها هشت ژن موکوس شناسایی شده است که پنج مورد آن پروتئینهای
ترشحی (Muc2، Muc5ac، Muc5b، Muc6 ovomucin) و سه مورد دیگر مولکولهایTransmembrane (Muc13، Muc16 وMuc1) هستند (Lang و همکاران، 2006).
موکوسهای ترشحی عمدتاً از سلولهای
گابلت آزاد میشوند. این سلولها بخشی از لایه اپیتلیال هستند که لومن روده را
از لامینا پروپریا جدا میکنند و لایه مخاطی تولید میکنند که عمدتاً از گلیکوپروتئینهای
موکوس تشکیل شده است. این موکوسها میتوانند از طریق ترشح (baseline secretion) یا وازیکولزدایی
ترکیبی
(compound exocytosis) آزاد
شوند (Kim و
Khan، 2013).
در مقابل، موکوسهای غشایی (Transmembrane mucins) بر
سطح انتروسیتها قرار دارند و نقش محافظتی مستقیم دارند (Kim و Khan، 2013).
موکوسها بهعنوان بخشی از لایه
مخاطی، از آسیب روده توسط محتویات لومن جلوگیری کرده و مانع چسبیدن پاتوژنها به
دیواره روده میشوند. ترکیب آنها میتواند تحت تأثیر مواد مغذی و ترکیبات
ضد میکروبی تغییر کند. برای مثال، مقدار موکوس خام (crude mucin) با افزایش
میزان ترئونین در جیره غذایی افزایش مییابد (Horn و
همکاران، 2009).
آزمایشهای in ovo
(اصطلاح “in ovo” به معنای داخل تخم است) نشان دادهاند که تجویز مانان
الیگوساکارید(MOS) سه روز قبل از هچ، سطح Muc2 را افزایش
میدهد
(Cheled-Shoval و همکاران، 2011). نتایج
مشابهی نیز در جوجههای گوشتی دریافتکننده جیره غذایی حاوی MOS مشاهده شده است؛ بهطوری که
بیان Muc2 در ژژونوم پرندگانی که MOS دریافت کردهاند، نسبت به پرندگان
بدون مکمل، بالاتر است.
برخی مطالعات به بررسی بیانmRNA موکوس پس
از مواجهه با NE پرداختهاند و تغییراتی در Muc2، Muc5ac و
Muc13 در روزهای پس از عفونت گزارش شده، اگرچه این نتایج همیشه
بین مطالعات یکسان نیست (Collier و همکاران، 2008). Collier و همکاران (2008) در روز پایانی عفونت و دو روز پس از
آن، افزایش بیان Muc2 mRNA
را در ایلئوم جوجههایی که با C. perfringens
و همعفونت با ایمریا مواجه شده بودند، مشاهده کردند.
Forder و همکاران (2012) افزایش بیان mRNA Muc5ac
را سه روز پس از مواجهه همزمان با NE گزارش کردند و در همان زمان
کاهش بیان Muc2 و Muc13 mRNA
مشاهده شد
Kitessa و همکاران (2014) نیز تغییراتی
در بیان Muc5ac و Muc13 در حضور عوامل مستعدکننده گزارش کردند، اما زمانی که C. perfringens به چالش
تجربی اضافه شد، سطح
mRNA مشابه گروه کنترل باقی ماند. این تفاوت ها ممکن است ناشی از
تفاوت در مدلهای آزمایشی باشد؛ زیرا پرندگان در روزهای متفاوتی تلقیح شدهاند،
همه مطالعات از همان عوامل مستعدکننده استفاده نکردهاند و علاوه بر آن، احتمال
تفاوت در بیان موکوس در مناطق مختلف روده باریک نیز وجود دارد.
سلولهای
اپیتلیال و ایمنی روده
پاتوژنهایی که میتوانند لایه
مخاطی را مختل کنند و از پپتیدهای ضد میکروبی فرار کنند، با اپیتلیوم روده
جوجه مواجه میشوند. سلولهای گابلت، انتروسیتها و لنفوسیتهای داخل اپیتلیال (IELs) بخشی از لایه اپیتلیال را تشکیل میدهند (Brisbin و همکاران،
2008). جمعیت
IEL شامل سلولهای NK، T و
B است (Gobel و همکاران، 2001).
رو به رو شدن با اپیتلیوم روده میتواند
گیرندههای شناسایی پاتوژن مانند Toll-like receptors (TLRs) را
فعال کند. این گیرندهها در انواع سلولها مانند سلولهای دندریتیک، هتروفیلها و
سلولهای اندوتلیال یافت میشوند. TLRها نقش مهمی در شناسایی الگوهای مولکولی مرتبط با
پاتوژن (PAMPs) دارند
که روی سطح باکتریها قرار دارند. فعال شدن این مولکولها مسیرهایی را تحریک میکند
که بیان مولکولهای التهابی را افزایش داده و باعث جذب سلولهای التهابی بیشتری
مانند سیتوکینها و کموکینها به محل عفونت میشوند (Kaiser، 2010).
ژنهای TLR در کل روده بیان میشوند که
نشاندهنده توانایی روده در شناسایی طیف گستردهای از پاتوژنهاست (Iqbal و همکاران، 2005). بیان ژنهای TLR در ایلئوم در چند روز اول پس از مواجهه با C. perfringens در روز ۱۸ زندگی تغییر میکند (Lu و
همکاران، 2009).
راهبردهای
کنترلی در برابر انتریت نکروتیک (NE)
اصول پیشگیری و کنترل NE عمدتاً بر پایه بیوسکیوریتی، بهداشت، مدیریت گله، استفاده از ضد
میکروبها و پیشگیری از عوامل مستعدکننده است (شکل ۵).

شکل ۵. راهبردهای پیشگیری از انتریت نکروتیک (NE)
نقش ضد
میکروبها
(Antimicrobials) در کنترل NE
ضد میکروبها در گذشته برای کنترل
بیماریها و افزایش بهرهوری به جیره غذایی یا آب طیور افزوده میشدند. این
ترکیبات میتوانند با کاهش تعداد پاتوژنهای فرصتطلب در میکروفلور روده،
خطر عفونتهای تحت بالینی را کاهش داده و هضم مواد مغذی را بهبود بخشند (Dibner وRichards ، 2005). اصطلاح ضد میکروب شامل مواد طبیعی، سنتزی و نیمهسنتزی است که برای مهار
رشد یا کشتن میکروارگانیسمها به کار میرود (Giguere، 2006).
مهمترین نگرانی در مورد
استفاده پیشگیرانه از آنتی بیوتیکها، انتقال مقاومت میکروبی به پاتوژنهای انسانی
از پاتوژنهای حیوانی است (سازمان جهانی بهداشت، 2001). علاوه بر این، محیطزیستگرایان
هشدار میدهند که فضولات طیور حاوی آرسنیک، فلزات سنگین و ضد میکروبها میتوانند
در محیط پخش شده و باعث گسترش ژنهای مقاومت میکروبی شوند (Osterberg و Wallinga ، 2004).
سوئد در سال
1986 نخستین کشور شد که استفاده از محرکهای رشد ضد میکروبی در دامپروری را ممنوع
کرد (Wierup، 2001). کشورهای
دیگر اتحادیه اروپا از جمله انگلیس و دانمارک نیز به دنبال این اقدام،
ممنوعیت مشابهی اعمال کردند (Wierup، 2001).
در دهه ۱۹۹۰،
استفاده از آوپارستین
(avoparcin) در جیره طیور باعث ایجاد ذخیرهای از انتروکوکهای
مقاوم به وانکومایسین
(VRE) در حیوانات شد و به همین دلیل، برخی کشورهای اتحادیه
اروپا استفاده از گروههای ضد میکروب مشابه با داروهای انسانی را در دامداریها
ممنوع کردند (Bates و همکاران، 1994).
در سال ۱۹۹۹،
اتحادیه اروپا تایلوزین، زینک باسیتراسین، ویرجینیامایسین، اسپیرامایسین و
آوپارستین را ممنوع کرد (Casewell و همکاران، 2003).
این ممنوعیت آنتیبیوتیکهای محرک
رشد باعث افزایش شیوع NE و به دنبال آن افزایش استفاده از آنتیبیوتیکهای
درمانی شد (Casewell
و همکاران، 2003).
پس از اجرای ممنوعیت آوپارستین، آلمان،
هلند و ایتالیا کاهش شیوع VRE در انسانها را گزارش کردند (Emborg و همکاران، 2001).
حساسیت C.
perfringens به آنتیبیوتیکها (Antimicrobial Susceptibility)
حساسیت C. perfringens
به آنتیبیوتیکها با روشهای مختلف تعیین شده است. برخی محققان
از حداقل غلظت مهاری
(MIC) برای توقف رشد ۵۰٪ یا ۹۰٪ از C. perfringens استفاده کرده و آن را به ترتیب MIC₅₀ و
MIC₉₀ نامیدهاند (Gharaibeh و همکاران، 2010؛ Slavic و همکاران، 2011). Gharaibeh و همکاران فعالیت هر آنتیبیوتیک روی جدایهها را با رتبهبندی MIC₅₀ و MIC₉₀ از کوچک به بزرگ توصیف کردند. روش دیگر استفاده از مقادیر
اپیدمیولوژیک حدی
(cutoff) بود که با ارزیابی توزیع دادههای MIC تعیین میشود (Johansson و
همکاران، 2004؛ Slavic و همکاران، 2011).
در بلژیک،
MIC جدایهها از خوک، گاو و مرغ بررسی شد. همه جدایهها به آوپارستین،
فورازولیدون، مونننسین، نیتروفوران، پنیسیلینG، رونیدازول
و تیامولین حساس بودند اما نسبت به فلاوومایسین
مقاوم بودند. جدایههای مرغ نسبت به کاربادوکس، کلرامفنیکل، اریترومایسین و
ویرجینیامایسین مقاوم بودند (Dutta و Devriese، 1980).
Watkins و همکاران (1997) بررسی کردند که ۴۸ جدایه C. perfringens از مرغ و
بوقلمون به چه میزان به آنتیبیوتیکها حساس هستند. در آمریکا، این جدایهها از ۲۶ مرغداری جوجه گوشتی و ۲۲ مرغداری بوقلمون جمعآوری شدند. آنها
گزارش کردند:
Johansson و همکاران (2004) حساسیت ۱۰۲ جدایه C.perfringens از ۸۹ مرغ گوشتی سالم یا بیمار، ۹ مرغ تخمگذار و ۴ بوقلمون را بررسی کردند. جدایهها از
سوئد، دانمارک و نروژ بین سالهای ۱۹۸۶ تا ۲۰۰۲ جمعآوری
شدند. همه جدایهها نسبت به آمپیسیلین، ناراسین، آویلامایسین، اریترومایسین و
وانکومایسین حساس بودند. همچنین ۳ درصد و ۱۵ درصد جدایهها از سوئد و دانمارک به
باسیتراسین مقاوم بودند و ۱۳ درصد جدایهها از نروژ نسبت به ویرجینیامایسین مقاوم
بودند.
Martel و همکاران (2004) مقاومت ۴۷ جدایه C. perfringens از ۳۱ مرغداری گوشتی در بلژیک را بررسی
کردند. همه جدایهها نسبت به مونینسین، لازالوسید، سالینومایسین، مادورامایسین،
ناراسین، آویلامایسین، تایلوزین و آموکسیسیلین حساس بودند، اما ۶۶ درصد جدایهها
مقاومت جزئی نسبت به کلرتتراسایکلین و اوکسیتتراسایکلین نشان دادند.
در یک مطالعه، Chalmers
و همکاران
(2008a) دریافتند که ۲۸ از ۶۱ جدایه
(۴۵.۹%) نسبت به باسیتراسین مقاوم هستند؛ ۱۷ از ۴۱ جدایه (۴۱.۵%) و ۱۱ از
۲۰ جدایه (۵۵%) از پرندگان بیمار و سالم جمعآوری شدند. در مطالعهای دیگر، Chalmers و
همکاران (2008b) گزارش کردند که ۳۹ از ۴۱ جدایه (۹۵.۱٪) مقاوم به باسیتراسین بودند؛
تمام جدایههای مقاوم از پرندگانی که باسیتراسین دریافت کرده بودند به دست آمده
بود و دو جدایه حساس از پرندگانی که باسیتراسین دریافت نکرده بودند. علت شیوع
بالای مقاومت به باسیتراسین ناشناخته بود، اما ممکن است ژنهای مقاومت بهصورت
افقی بین سویهها منتقل شده یا سویههای مقاوم نسبت به غیرمقاوم مزیت انتخابی
داشته باشند. همچنین
۱۶ از ۴۱ جدایه (۴۱.۴٪) نسبت به تتراسایکلین مقاوم بودند (با نقطه شکست ۴ میلیگرم/میلیلیتر طبق گفته Johansson و همکاران،
2004).
Gharaibeh و همکاران (2010) حساسیت آنتیبیوتیکی
کلستریدیوم پرفرنژانس را در ۱۵۵ مرغ گوشتی مبتلا به انتریت در اردن
بررسی کردند. نتایج
MIC نشان داد که حساسیت به آنتیبیوتیکها
متغیر است. کاهش حساسیت برخی آنتیبیوتیکها به مصرف آنها در واحدهای پرورش طیور
نسبت داده شد.
Gad و همکاران (2011) تعداد ۱۰۰ جدایه کلستریدیوم پرفرنژانس را که بین مارس ۲۰۰۸ تا مارس ۲۰۰۹ از گلههای بوقلمون در آلمان جمعآوری شده بود، از نظر
حساسیت آنتیبیوتیکی آزمایش کردند. آنتیبیوتیکهای بتالاکتام و همچنین ترکیب
لینکومایسین، اسپکتینومایسین و تایلوزین بر همه جدایهها مؤثر بودند. بیشتر جدایهها
به انروفلوکساسین (۹۸٪)، اگزاسیلین (۸۳٪)، تیامولین (۸۰٪)، تیلمیکوزین (۸۰٪) و تریمتوپریم/سولفامتوکسازول (۷۲٪) حساس بودند. اما بیشتر جدایهها نسبت به اسپکتینومایسین (۷۴٪)، نئومایسین (۹۴٪) و کولیستین (۱۰۰٪) مقاوم بودند.
Slavic
و همکاران (2011)
MIC صد جدایه کلستریدیوم پرفرنژانس بهدستآمده
از مرغهای گوشتی در انتاریو را بررسی کردند. نتایج نشان داد مقاومت به باسیتراسین
(۶۴٪)، ویرجینیامایسین (۲۵٪)، تتراسایکلین (۶۲٪)، اریترومایسین (۲٪)، کلیندامایسین (۲٪) و مترونیدازول (۱٪). هیچ مقاومتی نسبت به سالینومایسین (۰٪) و فلورفنیکل (۰٪) گزارش نشد. آنها پیشنهاد کردند که الگویی از افزایش مقاومت
کلستریدیوم پرفرنژانس نسبت به برخی آنتیبیوتیکهای رایج در پیشگیری و درمان
بیماریها وجود دارد. کاهش حساسیت نسبت به چندین آنتیبیوتیک گزارش شد.
Gad و همکاران (2012) تعداد ۴۶ جدایه کلستریدیوم پرفرنژانس را که بین سالهای ۲۰۰۸ و ۲۰۰۹ از گلههای مرغ تخمگذار صنعتی جمعآوری شده بود، از
نظر مقاومت به ۱۶ آنتیبیوتیک آزمایش کردند. آنتیبیوتیکهای بتالاکتام،
تایلوزین، داکسیسایکلین، تتراسایکلین، انروفلوکساسین، تریمتوپریم/سولفامتوکسازول،
لینکومایسین و تیلمیکوزین بر همه جدایهها مؤثر بودند. مقاومت نسبت به
اریترومایسین (۱۷.۴٪) و تیامولین (۱۹.۶٪) مشاهده شد.
راهکارهای
جایگزین برای کنترل نکروتیک انتریت (NE)
گزینههای مختلفی بهعنوان
جایگزین آنتیبیوتیکها در خوراک پیشنهاد شدهاند
Caly .و همکاران (2015) مروری بر راهبردهای مختلف کنترل کلستریدیوم پرفرنژانس ارائه
کردهاند. به طور کلی، عفونتهای میکروبی با کلونیزه کردن دستگاه گوارش، بر وزن
نهایی بدن، سلامت روده و کیفیت گوشت جوجههای گوشتی تأثیر منفی گذاشته و در نهایت
بهرهوری پرورش طیور را کاهش میدهند (Abd El-Hack و همکاران، 2020a).
آنتیبیوتیکها بهطور مؤثر از رشد و تکثیر میکروارگانیسمها
جلوگیری میکنند، اما مشکل زمانی آغاز میشود که باکتریهای مقاوم به آنتیبیوتیک
ظاهر شوند. استفاده از ترکیبات جایگزین سازگار با محیطزیست، میتواند این مشکل را
برطرف سازد (Abd El-Hack و
همکاران، 2020b,c؛ Abo Ghanima و همکاران، 2021). این جایگزینها شامل عصارههای گیاهی (El-Saadony و
همکاران، 2021b)، افزودنیهای فیتوژنیک خوراک
(Abdelnour و همکاران، 2020a,b؛ Ashour و
همکاران، 2021؛ Raza و همکاران، 2021؛ Seidavi
و همکاران، 2021a,b)، آنتیبادیهای تخممرغی (Wilkie و همکاران، 2006)، آنزیمهای خوراکی (Llamas-Moya و
همکاران، 2020)، واکسیناسیون (Kulkarni و همکاران، 2007)، و همچنین اصلاح جیره از طریق
انتخاب مواد اولیه، نوع غلات، فرآوری خوراک و سطح پروتئین جیره هستند (Caly و
همکاران، 2015).
پروبیوتیکها بهعنوان «مکمل غذایی میکروبی زنده که با بهبود تعادل
میکروبی روده بر میزبان اثر مفید دارد» تعریف میشوند
(Fuller، 1999). پروبیوتیکها نقش مهمی در
متابولیسم، تحریک سیستم ایمنی و پیشگیری از بیماریها دارند
(Edens، 2003). همچنین میتوانند با تعدیل
میکروبیوتای روده، ضریب تبدیل خوراک، قابلیت هضم و جذب مواد مغذی
را بهبود دهند (Kan
و همکاران، 2021).
علاوه بر این، مطالعات نشان
دادهاند که پروبیوتیکها از بروز بیماریهای مختلف گوارشی در تولیدات دامی
جلوگیری میکنند (La Ragione و همکاران، 2004).
بنابراین یکی از گزینههای جایگزین
مناسب برای آنتیبیوتیکها محسوب میشوند. پروبیوتیکها را میتوان از طریق آب
آشامیدنی، خوراک یا اسپری به پرندگان داد. هرچند مکانیسم دقیق عملکرد آنها کاملاً
روشن نیست، اما احتمالاً به دو روش عمل میکنند:
پروبیوتیکها همچنین نشان دادهاند
که میتوانند مورفولوژی سلولهای رودهای را تغییر دهند؛ از جمله بزرگتر شدن هسته
سلولها و افزایش فعالیت میتوزی (Kabir، 2009).
Chichlowski و همکاران (2007) مشاهده کردند که افزودن Bacillus subtilisبه خوراک مرغ موجب افزایش طول پرزهای روده و تقسیم سلولی
میشود. پرمصرفترین پروبیوتیکها برای طیور شامل گونههای Bacillus، Aspergillus، Lactobacillus، Bifidobacterium، Candida و Streptomyces هستند. در این میان، Lactobacillus و Bacillus بیشترین
کاربرد را علیه نکروتیک انتریت ناشی از کلستریدیوم پرفرنژانس دارند (Jadamus و
همکاران، 2001). پرندگانی که پروبیوتیک دریافت کرده بودند، تیتر آنتیبادی
بالاتری نسبت به گروه کنترل داشتند. همچنین، پروبیوتیکها میتوانند با تولید
باکتریوسینها (مواد ضدباکتری) و مهار تولید سموم باکتریایی از عفونت جلوگیری
کنند. برای مثال، Bacillus subtilis PB6 توانسته است پرندگان را در برابر NE ناشی از کلستریدیوم
پرفرنژانس محافظت کرده و سلامت روده را بهبود دهد
(Jayaraman و همکاران، 2013).
میکروبیوتا
و متابولیتهای آن
میکروبیوتا میتواند در پیشگیری
و درمان بیماریهای مختلف نقش داشته باشد. انتقال میکروبیوتا از پرندگان بالغ سالم
به جوجههای دو روزه، توانسته است آنها را در برابر کلونیزاسیون سکومی سالمونلا
اینفانتیس محافظت کند (Rantala و
Nurmi، 1973).
در بیماریهای گوارشی انسان نیز، مانند
عفونت Clostridium difficile، انتقال میکروبیوتا به بدن بیمار درمان موفقی به همراه داشته است (Lawley و
همکاران، 2012).
میکروبیوتا از طریق مکانیسمهای
مختلفی مانع بروز یا پیشرفت بیماریها در حیوانات میشود. یکی از این مکانیسمها حذف
رقابتی (Competitive Exclusion, CE) است که طی آن میکروبیوتا باکتریهای بیماریزا را در
رقابت برای مواد مغذی یا جایگاههای اتصال (کلونیزاسیون) کنار میزند. روی یکی از مواد معدنی حیاتی برای هر دو گروه، یعنی
میکروبیوتای مفید و باکتریهای پاتوژن، به شمار میآید. میکروبیوتای سالم در روده
با رقابت برای مواد مغذی، از کلونیزاسیون باکتریهای بیماریزا در جوجههای جوان
جلوگیری میکند (Schneitz، 2005).
مکانیسم دیگر در حذف رقابتی، تولید
باکتریوسین است. باکتریوسین ترشحشده از میکروبیوتا خاصیت باکتریکشی دارد و
بنابراین میتواند در پیشگیری و درمان عفونتهای ناشی از باکتریهای بیماریزا به
کار رود (Caly
و همکاران، 2015).
دستگاه گوارش میزبان ــ از جمله اپیتلیوم
روده و سیستم ایمنی ــ بخشی از رشد و بلوغ خود را مدیون فعالیت میکروبیوتای روده
است؛ از طریق تشکیل اندامهای لنفوئیدی، تولید پپتیدهای آنتی بیوتیکی و ایمونوگلوبولین
A، و همچنین
فعالسازی و تمایز لنفوسیتها.
Forder
و همکاران (2007) اثر میکروبیوتا بر ساختار سلولهای گابلت و مخاطی
پس از خروج از تخم را نشان دادند. در مقایسه با حیوانات عاری از میکروب، حیواناتی
که بهطور متعارف پرورش یافتند، تکامل بهتری در مورفولوژی روده شامل لایه مخاطی،
لایه اپیتلیال، لامینا پروپریا، طول پرزها و عمق کریپت داشتند(Deplancke و Gaskins، 2001).
افزون بر این، باکتریهای مفید تأثیر
چشمگیری بر ذخیره T سلولها و بیان سیتوکینها در روده نشان دادهاند (Mwangi و
همکاران، 2010). میکروبیوتای روده نقشی اساسی در رشد و کارایی حیوان
دارد. هرچند هنوز بحثبرانگیز است که آیا میکروبیوتا میتواند پیش از خروج از تخم
وارد جنین شود یا خیر، مطالعات زیادی نشان دادهاند که خوراک، آب، شرایط جوجهکشی
و حملونقل بر ترکیب و کلونیزاسیون میکروبیوتای جوجههای پس از هچ اثرگذارند (Kers و
همکاران، 2018).
میکروبیوتای روده و متابولیتهای
آنها هضم، جذب و متابولیسم مواد مغذی را بهبود بخشیده و در نتیجه بهرهوری پرورش
جوجههای گوشتی را ارتقا میدهند. این اثر عمدتاً به تولید اسیدهای چرب کوتاهزنجیر (SCFAs) شامل استات، پروپیونات، بوتیرات و لاکتات، بههمراه اسیدهای آمینه و
ویتامینهای B و K نسبت داده میشود (Yadav و Jha، 2019). این اسیدهای چرب کوتاهزنجیر از تخمیر پلیساکاریدها و
ترکیبات غیرقابلهضم (NSP) حاصل میشوند و با تأمین انرژی برای سلولهای اپیتلیال
روده، موجب افزایش ارتفاع پرزها و بهبود جذب میگردند
(De
Vadder و همکاران، 2014). این یافته توسط مطالعاتی بر روی
موشهای عاری از میکروب و موشهای معمولی تأیید شده است؛ موشهای عاری از میکروب
نسبت به موشهای معمولی پرزهای کوتاهتر، سطح روده کمتر و رشد ناقص میکروویلی
داشتند (Deplancke
و Gaskins، 2001). افزون بر این، SCFAها در تنظیم گلوکز، متابولیسم انرژی و چربی نیز نقش ایفا
میکنند (Den Besten
و همکاران، 2013).
ترکیبات
فیتوژنیک مورد استفاده در کنترل انتریت
نکروتیک القاشده با C. perfringens
امروزه جایگزینهای مؤثر زیادی
برای آنتیبیوتیکها وجود دارد (جدول 1) که در کنترل انتریت نکروتیک کاربرد دارند؛
از جمله عصارههای گیاهی (Abou-Kassem و همکاران، 2021؛ Reda و همکاران، 2021a)، پپتیدهای زیستفعال
(El-Saadony و همکاران، 2021b؛ Saad و همکاران، 2021c)، افزودنیهای گیاهی خوراک در جیره طیور (Ashour و
همکاران، 2020a,b)، نانوذرات سنتزشده زیستی
(Reda و همکاران، 2020؛ Sheiha و همکاران، 2020؛ Abd
El-Ghany و همکاران، 2021)، پروبیوتیکها (Abd El-Hack و
همکاران، 2021c)، پریبیوتیکها، سینبیوتیکها و آنزیمها
(Llamas-Moya و همکاران، 2020).
اثر بخشی بسیاری از ترکیبات
فیتوژنیک در کاهش بروز و شدت انتریت نکروتیک مورد بررسی قرار گرفته است (جدول 1).
بهعنوان نمونه:
همگی توانستهاند بهطور موفق
از رشد C. perfringens
جلوگیری کرده و کارایی پرورش جوجههای گوشتی را بهبود بخشند (Gharaibeh و
همکاران، 2021).
ﺟﺪﻭﻝ 1: ﺗﺄﺛﯿﺮ ﻋﺼﺎﺭﻩ ﻫﺎ ﻭ ﺭﻭﻏﻦ ﻫﺎﯼ
ﺿﺮﻭﺭﯼ ﮔﯿﺎﻫﺎﻥ ﻣﺨﺘﻠﻒ ﺑﺮ ﮐﻠﺴﺘﺮﯾﺪﯾﻮﻡ ﭘﺮﻓﺮﯾﻨﺠﻨﺲ عامل ﺍﯾﺠﺎﺩ ﺍﻧﺘﺮﯾﺖ ﻧﮑﺮﻭﺗﯿﮏ (NE)
|
نوع گیاه و روغنهای
اسانسی |
دوز توصیه شده |
الگوی بیماری |
اثر اصلی |
منابع |
|
پودر زردچوبه (Curcuma longa) |
2 کیلوگرم در هر تن خوراک |
بدون عفونت آزمایشی |
بهطور معنیداری بار C. perfringens و مرگومیر جوجهها را کاهش میدهد. |
ﻋﻠﯽﻭ ﻫﻤﮑﺎﺭﺍﻥ، ۲۰۲۰ ﮐﺎﻟﯿﮏ ﻭ ﻫﻤﮑﺎﺭﺍﻥ، ۲۰۱۹ |
|
عصاره یوکا موجاوه (Yucca schidigera) |
۵۶۷ گرم در هر تن برای جیره آغازین،
۴۵۴ گرم در هر تن برای جیره رشد، و ۳۴۰ گرم در هر تن برای
جیره پایانی |
مدل
انتریت نکروتیک طبیعی با چالش غیرمستقیم از طریق واکسن Coccivac- B52 در روز ۷
|
در
طول فاز چالش اولیه انتریت نکروتیک، عملکرد جوجههای گوشتی را بهطور معنیداری
بهبود میبخشد و همچنین در دوره رشد نیز مؤثر است، اما تأثیری بر ضایعات انتریت
نکروتیک در روده کوچک ندارد. |
کالیگ و
همکاران، 2019 |
|
انگور ماسکاداین (Vitis rotundifolia) |
۵، ۲۰ و
۵۰ کیلوگرم در هر تن خوراک |
۲ × ۱۰7 CFU/mL در روزهای ۱۹، ۲۰ و ۲۱، که پیش از آن در روز ۱۴ با Eimeria acervulina و Eimeria maxima همراه بوده است.
|
عصارهها در ۵ و ۲۰ کیلوگرم در هر تن خوراک ،
پس از چالش با C. perfringens موجب بهبود افزایش
وزن بدن و کاهش مرگومیر و ضایعات پا میشوند. |
مکدونالد و همکاران 2018 |
|
پونه کوهی (Origanum vulgare) |
100
PPM |
بدون
عفونت آزمایشی |
تعداد
C.
perfringens در دستگاه گوارش و مدفوع را کاهش میدهد. |
Mitch, 2004 |
|
آویشن (Thymus vulgaris) |
۴ گرم
در هر تن |
۱ × ۱۰⁹ CFU از C. perfringens به ازای هر پرنده در روز ۱۸، که پیش از آن در
روز ۱۴ با Eimeria maxima بهصورت
خوراکی آلوده شده است. |
افزایش وزن بدن را به همراه دارد و پاسخ التهابی مرتبط
با انتریت نکروتیک و نمره ضایعات ماکروسکوپی روده را کاهش میدهد. |
Lee et al., 2013 |
|
مریم گلی شیرین (Artemisia annua) |
10 کیلوگرم در هر تن خوراک |
تجویز
جیرههای شامل آرد ماهی همراه با ۲۰۰ میلیلیتر از ۱۰⁶ C. perfringens در
روزهای ۱۷، ۱۸، ۱۹ و ۲۰، همراه با تجویز دوز ۱۰ برابری واکسن Paracox-5.
|
افزایش
وزن بدن و مصرف خوراک را کاهش میدهد، اما ضریب تبدیل خوراک را بهبود میبخشد. |
Engberg et al., 2012 |
|
ترکیبی از تیمول، سینامآلدئید و اکالیپتوس |
150 گرم در تن |
۴.۱۰⁸ CFU از C. perfringens در روزهای ۱۹، ۲۰، ۲۱ و ۲۲ (۳ بار در روز)، با استفاده از جیره مبتنی بر
گندم/چاودار (۴۳٪/۷.۵٪) و با منبع پروتئین
شامل آرد سویا و آرد ماهی (۳۰٪)، که در ادامه در
روز ۲۰ دوز ۱۰ برابری واکسن Paracox-5 داده شده است. |
نمره ضایعات ماکروسکوپی روده القا شده توسط C. perfringens را کاهش میدهد. |
Timbermont et al., 2010 |
|
روغن زنجبیل و کارواکرول ۱٪ |
5/1 کیلوگرم در هر تن خوراک |
۲ میلیلیتر سوسپانسیون C. perfringens نوع A (۶–۸ × ۱۰⁸ CFU) بهصورت خوراکی در روزهای ۱۸،
۱۹، ۲۰ و ۲۱، سه بار در روز (۰۸:۰۰، ۱۲:۰۰
و ۱۶:۰۰)، که پیش از آن در روز ۱۶ واکسن بیماری بورس
از طریق آب آشامیدنی جهت القای نقص ایمنی داده شده و در ادامه در روز ۱۹
دوز ۱۰ برابری واکسن Paracox-5 تجویز شده است. |
عملکرد
رشد را بهبود میبخشد، نمره ضایعات ماکروسکوپی و میکروسکوپی روده را کاهش میدهد،
و طول پرزهای اپیتلیال و نسبت پرز به کریپت را افزایش میدهد.
|
Jerzcele et al., 2012 |
|
روغن نعناع فلفلی و میکروامولسیون روغن نعناع فلفلی (۱۵٪
روغن/آب) |
روغن نعناع فلفلی (۰.۵ میلیلیتر در میلیلیتر
آب) و میکروامولسیون آن (۰.۲۵ میلیلیتر در میلیلیتر آب)
|
۱ میلیلیتر ) ۱ × ۱۰⁸
CFU (C.
perfringens بهصورت ۳
دوز متوالی در روزهای ۱۴، ۱۵ و ۱۶
|
باعث کاهش ضایعات ماکروسکوپی و مرگومیر میشود، تعداد
واحدهای کلنی (CFU) را کاهش داده و
عملکرد رشد را بهبود میبخشد.
|
Sorour
et al., 2021 |
|
پونه کوهی (Origanum vulgare زیرگونه hirtum) |
10
کیلوگرم در هر تن خوراک |
بدون
عفونت آزمایشی |
عملکرد
رشد را بهبود میبخشد. |
Cross et al., 2007 |
|
رزماری (Rosmarinus officinalis) |
10 کیلوگرم در هر تن خوراک |
بدون عفونت آزمایشی |
تعداد باکتریهای C. perfringens در سکوم و مدفوع را کاهش میدهد و تأثیری بر باکتریهای
اسید لاکتیک ندارد. |
Cross et al., 2007 |
|
روغنهای اسانسی و اسیدهای آلی |
500
گرم در هر تن خوراک |
۲.۲ × ۱۰⁸ CFU در روز بهصورت خوراکی در
روزهای ۱۸ تا ۲۰، که پیش از آن در روز ۱۴ با Eimeria maxima و Eimeria necatrix همراه
بوده است. |
ضریب
تبدیل خوراک را بهبود میبخشد، ارتفاع پرزها و نسبت پرز به کریپت را افزایش میدهد،
تعداد C. perfringens روده، انتقال C. perfringens به کبد، نمرات ضایعات روده و میزان فلورسانس سرمی ایزوتیوسیانات
دکسترن را کاهش میدهد. |
Pham et al., 2021 |
|
بابونه وحشی یا یارو (Achillea millefolium
واریته alba) |
10 کیلوگرم در هر تن خوراک |
بدون عفونت آزمایشی |
|
Cross et al., 2007 |
واکسیناسیون
برای انتریت نکروتیک (NE)
با وجود اینکه واکسیناسیون یک
گزینه جایگزین برای داروهای آنتی بیوتیکی در کنترل بیماری است، ایمنی در برابر NE هنوز بهطور کامل شناخته نشده
است (Kulkarni و
همکاران، ۲۰۰۷). آلفاتوکسین، که ایمنزاترین پروتئین تولید شده توسط C.
perfringens است،
سیستم ایمنی را تحریک کرده و موجب تولید آنتیبادی میشود (Kulkarni و
همکاران، ۲۰۰۷). در مقایسه جوجههای سالم با جوجههای مبتلا به NE، جوجههای سالم سطح بالاتری از آنتیبادیها (IgY) علیه هر دو NetB و α-توکسین نشان دادند (Lee و همکاران، ۲۰۱۲). این یافته نشاندهنده اهمیت این
آنتیبادیها در محافظت از جوجههای گوشتی در برابر NE است (Lee و همکاران، ۲۰۱۲).
مطالعات متعددی برای پیشگیری از NE با استفاده از واکسن انجام شده
است. برخی از این واکسنها شامل: واکسنهای توکسوئیدی (سم غیرفعال)، واکسنهای
زنده تخفیف حدت یافته. این واکسنها میتوانند از طریق خوراک، آب، اسپری در
جوجهکشی و تزریق به داخل تخممرغ in ovo تجویز شوند
(Mot
و همکاران، 2019).
Keyburn
و همکاران (۲۰۱۳b) جوجههای گوشتی را بهصورت زیرجلدی با NetB نوترکیب
خالص (rNetB)، باکتریهای کشته شده با فرمالین و توکسوئید بدون سلول یا همراه
با مکمل rNetB ایمنسازی کردند. آنها دریافتند که واکسیناسیون تنها با NetB پرندگان را در
شرایط مزرعه در برابر NE محافظت نمیکند، اما میتواند هنگامی که با سایر آنتیژنهای سلولی
ترکیب شود، از جوجهها محافظت کند. ایمنسازی مادران با واکسن توکسوئیدی تقویتشده
با NetB یک روش مفید
برای کنترل NE در جوجهها است (Keyburn و همکاران، ۲۰۱۳a).
Mishra
و Smyth (۲۰۱۷) یک واکسن خوراکی برای جوجههای گوشتی علیه NE توسعه دادند که از سویه غیرموذی
مثبت NetB نوع A C. perfringens استفاده میکند و این واکسن ایمنی حفاظتی را در گروه
واکسینهشده نسبت به گروه کنترل ایجاد کرد.
واکسیناسیون با توکسین تغییر
شکل یافته یا سایر پروتئینهای ایمنزای ترشحشده به نظر میرسد روش منطقی برای
دفاع در برابر باکتریهای تولیدکننده توکسین باشد. ایمنسازی نیازمند ترکیبی از
آنتیژنهای ایمنزا و پروتئینهای حفاظتی مختلف و برنامههای چنددوزی است که در
صنعت جوجهکشی قابل اجرا نیست، زیرا واکسیناسیون تنها برای جوجههای یکروزه
معمولاً محافظتکننده نیست. یک واکسن غیرفعال شامل توکسوئید α-توکسین نوع A C. perfringens همراه با افزودنی روغنی در جوجهها (دو دوز در سن ۱۰–۱۴ روز و ۴–۱۰ هفته بعد)
برای ایجاد حفاظت غیرفعال در جوجهها علیه NE استفاده شد (Mot و همکاران، ۲۰۱۴). این واکسن در برابر چالش شدید
دادهشده در خوراک محافظتی ایجاد نکرد. مطالعات نشان دادهاند که NE میتواند با واکسنهای زنده تخفیف
حدت یافته کنترل شود (Jiang و
همکاران، ۲۰۱۵). واکسنهای نوترکیب
زنده تخفیف حدت یافتهSalmonella
که بهصورت خوراکی به جوجهها داده میشوند، حفاظت مقرونبهصرفهای در
برابر C. perfringens
ارائه میدهند (Jiang و همکاران، ۲۰۱۵).
با وجود مطالعاتی که ایمنی جزئی
توسط این واکسنها نشان دادهاند، هنوز هیچ واکسن تجاری مؤثری علیه NE القا شده توسط C. perfringens وجود ندارد.
علاوه بر این، توسعه آینده واکسن باید حفاظت در برابر NE و توکسینها را تأیید کند، عملکرد پرنده را بهبود بخشد و
روشهای ارائه واکسن به جوجههای گوشتی را بهینهسازی نماید.
نتیجهگیری
عفونت انتریت نکروتیک (NE) تهدیدی جدی برای صنعت طیور محسوب میشود که میتواند خسارات اقتصادی
شدیدی به همراه داشته باشد. پس از ممنوعیت جهانی استفاده از آنتیبیوتیکها، نیاز
فوری به یافتن جایگزینهای مؤثر برای کنترل این بیماری وجود دارد.
پروبیوتیکها، پریبیوتیکها،
سینبیوتیکها، روغنهای اسانسی، عصارههای گیاهی و واکسنهای حفاظتی، جایگزینهای
مؤثری برای آنتیبیوتیکها هستند که میتوانند همراه با رعایت اقدامات بهداشتی و
امنیت زیستی، اثرات منفی NE در طیور را کاهش دهند.
دکتر پرستو شکری

کنسانتره طیور

کنسانتره طیور

کنسانتره طیور

مکمل ویتامینه – معدنی

مکمل ویتامینه – معدنی

مکمل ویتامینه – معدنی

سایر محصولات

سایر محصولات